 Back
BackDNA Replication, Repair, and Recombination

 14:26
14:26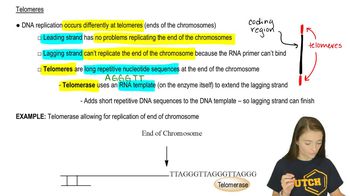

Terms in this set (29)
Semiconservative replication means each new DNA molecule contains one original strand and one newly synthesized strand.
DNA replication begins at specific sequences called replication origins where initiation proteins bind.
DNA polymerase synthesizes new DNA strands in the 5’ to 3’ direction, adding nucleotides to the 3’ end.
The leading strand is synthesized continuously, while the lagging strand is synthesized discontinuously as Okazaki fragments.
RNA primers (~10 nucleotides) are synthesized by primase to provide a starting point for DNA polymerase.
Repair polymerase replaces RNA primers with DNA nucleotides during replication.
DNA ligase joins Okazaki fragments together to form a continuous lagging strand.
DNA helicases unwind the double helix by breaking hydrogen bonds between base pairs.
SSB proteins bind single-stranded DNA to prevent it from re-forming a double helix during replication.
DNA topoisomerases prevent supercoiling by cutting and rejoining DNA strands ahead of the replication fork.
The sliding clamp keeps DNA polymerase attached to the DNA strand for efficient replication.
The lagging strand cannot fully replicate chromosome ends because RNA primers cannot bind there, requiring telomerase.
Telomerase extends the lagging strand by adding repetitive DNA sequences using its RNA template.
DNA replication has an error rate of about 1 in 10 million nucleotides due to proofreading.
Proofreading is DNA polymerase's ability to remove mismatched bases using its 3’ to 5’ exonuclease activity before continuing synthesis.
Energy from hydrolysis of two phosphate groups from dNTPs powers DNA synthesis at the 3’ end of the growing strand.
Depurination is the spontaneous loss of purine bases (A or G) from DNA, creating abasic sites.
Deamination chemically converts one base into another, e.g., cytosine to uracil.
UV light causes adjacent thymine bases to form covalent dimers, distorting DNA.
Key repair mechanisms include mismatch repair, base excision repair, nucleotide excision repair, and double strand break repair.
Mismatch repair fixes incorrectly paired bases that distort the DNA helix.
BER removes damaged bases like uracil using DNA glycosylase and replaces them with correct bases.
NER removes bulky lesions like thymine dimers by cutting out damaged DNA and sealing the gap with DNA ligase.
Nonhomologous end joining directly joins broken ends; homologous recombination uses an undamaged template for accurate repair.
A double strand break occurs, initiating repair using a homologous DNA template.
RecA binds single-stranded broken and undamaged DNA to facilitate strand invasion and pairing.
Holliday junctions are cross-shaped DNA structures formed during homologous recombination connecting four DNA strands.
Branch migration moves the Holliday junction along DNA, increasing the region available for repair.
Cleavage can cause crossing-over leading to DNA exchange or non-complementary regions that may be repaired later.